





Competition is one of the major types of interactions between species in nature. It is especially important in biodiverse communities such as tropical forests, where a large number of plant species must compete for a small number of resources, such as water, light, and nutrients. Yet, we still know little about how competitive interactions influence what kinds of species can coexist.
Considering that other forces such as dispersal and chance events are known to affect ecological dynamics, one pressing question in community ecology is whether competition leaves a recognizable and detectable signal in species assemblages. For example, can we say anything general about what characteristics provide species with a competitive advantage over others such that they become abundant in their community or can invade existing communities?
Species phenotypic traits mediate competition. For example, two species with similar traits are likely to have similar resource needs and similar strategies for acquiring them and, therefore, compete more strongly with each other than two species with dissimilar traits (Figure 1A). It follows that communities where competition is a driving force may display specific patterns in the phenotypic composition of their species. Imagine a community with three species that cannot coexist because they are too mutually similar. The species with the intermediate trait will be under the strongest competitive pressure and eventually will be competitively excluded. Intuitively, this should lead to communities where species are more mutually dissimilar than expected by chance. Indeed, this idea of limiting similarity among competing species is a classical principle in community ecology, dating back to the 1950s. However, it has found limited empirical support, and in fact, more recent theoretical research has shown that it will only arise in special circumstances.
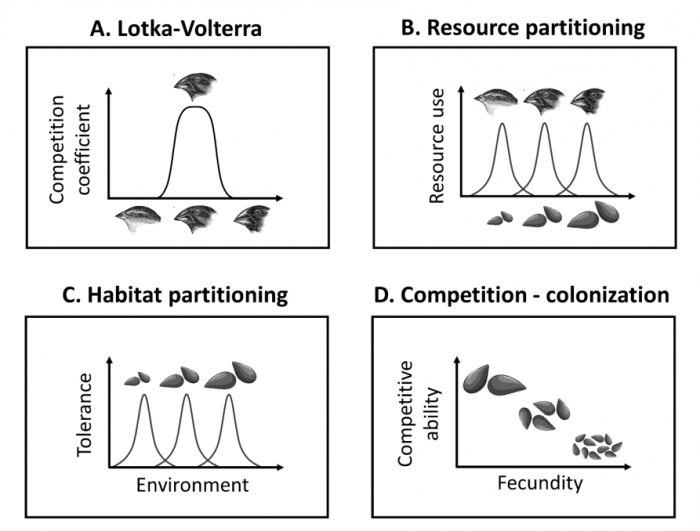
Figure 1. Schematic representation of competition models. A. Lotka-Volterra: competition is directly tied to similarity in species traits, here beak size in birds. The curve shows competitive impact by each species on the focal species, increasing with trait similarity. B. Resource partitioning: we assume a continuum of substitutable resources (seeds of different sizes), and species traits (beak size) determine their use of the different resources. C. Habitat partitioning: species with different traits (seed size) are optimally adapted to different environments (for example, soil pH). D. Competition-colonization tradeoff: depending on seed size, species trade-off fecundity (seed output) with the ability to win sites against other species. Image republished with permission from PLOS from https://doi.org/10.1371/journal.pcbi.1006688
In the three-species example above, it turns out that the more similar the intermediate species is to one of the other two, the longer it takes to be excluded. This may sound counterintuitive, and yet it has major consequences for how competitive exclusion plays out in communities with many species. First, a set of species whose traits are spaced enough from one another to stably coexist gets an advantage in the competitive sorting process, either because of initial conditions or because they are favored by the environment. Once these favored traits emerge, all other species are competitively excluded at a rate inversely related to their trait separation from the closest favored trait. This results in a transient pattern of distinct clusters of species with similar ecological strategies. Limiting similarity will only occur once all weaker species have been fully excluded, which may take an indefinitely long time.
In our study, we used simulations of competitive dynamics models to show that trait-based clustering is a robust signal of community assembly under competition, going beyond the simplified scenarios where it was previously known to occur.
We showed that dispersal of individuals into the community via immigration from surrounding communities turns these transient clusters into permanent features, as population losses to competition are counterbalanced by an inflow of new individuals. Further, we found that chance events known as demographic stochasticity will add noise to the pattern, but the clusters will typically still be detectable with appropriate statistical tools (Figure 2). The pattern is strongest when diversity is high – i.e. when there are many more species than clusters – and when immigration is intermediate – neither so low that most species are fully excluded between immigration events nor so high that it overcomes competition as the main process shaping the community.
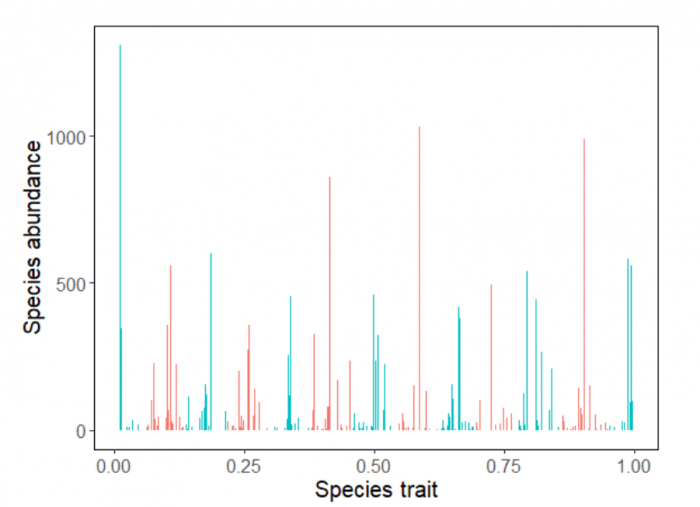
Figure 2. Trait-based clustering in a species assemblage. A community of 232 species totaling 21,000 individuals, generated via simulation of a competitive dynamics model. Our statistical analysis reveals trait-based clustering among species, with 13 color-coded clusters in this example, where each species is represented by one stem. Image republished with permission from PLOS from https://doi.org/10.1371/journal.pcbi.1006688
To check that clusters emerge generally under competition, we analyzed models describing different types of competitive dynamics. In addition to the phenomenological model described above where competitors directly affect each other via trait similarity (Figure 1A), we considered competition for substitutable resources where species differ by preferred resource (e.g. animals with overlapping but different dietary preferences, Figure 1B), competition for recruitment where species differ by optimal habitat (e.g. plants optimally adapted to low vs high soil pH, Figure 1C), and competition for recruitment under a trade-off between seed production and seedling survival, known as a competition-colonization tradeoff (Figure 1D).
We found that clustering occurred generally across these distinct types of competition. The strength of the pattern varied across different scenarios: for example, under resource partitioning, clustering was stronger when consumers were able to substantially deplete resource populations because resource depletion caused species to cluster around the remaining resources. However, the key result is that clustering is a robust outcome of competitive dynamics, tending to emerge when simple and intuitive connections between competition and trait similarity hold.
While cluster emergence under competition was general, the characteristics of the clusters varied: both the shape of the clusters – i.e. the relationship between species abundance and trait separation from the cluster center – and the trait separation between cluster centers were highly idiosyncratic across competition types and scenarios. This variation in shape and scale suggests that one is more likely to succeed in detecting the pattern by focusing on general aspects of clustering than on the particular shape or distribution of the clusters. Moreover, a single one-size-fits-all tool for detecting clusters across species assemblages may not exist. Indeed, in our study, we observed that different methods were best suited for different competition scenarios.
Clustering as a signature of coexistence under competition is an update to the prevalent paradigm that competing species will display greater-than-chance mutual differences. Our finding that clusters appear under various types of competition and can be easily maintained by immigration, even when confounding stochastic forces are at play, suggests that clusters are a likely feature of nature beyond the instances where it is currently known to occur. For example, in tropical forests, where both competition and dispersal are recognized as key drivers of community assembly, clustering could help explain the observed high diversity and seemingly continuous phenotypic variation. In fact, our preliminary results applying our methods to forest data indicate that tropical forests may be structured into distinct groups of species clustered by maximum height. If confirmed, these results will support the hypothesis that competition for light is a critical force in these forests, and show that competitive dynamics actually shape the community into recognizable phenotypic patterns.
These findings are described in the article entitled Generalizing clusters of similar species as a signature of coexistence under competition, recently published in the journal PLOS Computational Biology.
Related Posts
 Duck-Billed Dinosaurs Uncovered In Aniakchak, Alaska
Duck-Billed Dinosaurs Uncovered In Aniakchak, Alaska Cryptic Diversity In Vietnam’s Limestone Karst Habitats
Cryptic Diversity In Vietnam’s Limestone Karst Habitats An Improved Method To Remove Debris From Cyst Nematode Egg Suspensions And Computer-Aided Technologies For Egg Counting
An Improved Method To Remove Debris From Cyst Nematode Egg Suspensions And Computer-Aided Technologies For Egg Counting The Footprints Of Urbanization, Industrialization, And Agriculture On River Beds: Heavy Metal Contamination Assessment And Source Identification In River Sediments In Eastern China
The Footprints Of Urbanization, Industrialization, And Agriculture On River Beds: Heavy Metal Contamination Assessment And Source Identification In River Sediments In Eastern China Aging Dolphins Via Pectoral Flipper Radiography
Aging Dolphins Via Pectoral Flipper Radiography Glycoalkaloids In Potatoes: The Effect Of Biostimulants And Herbicides
Glycoalkaloids In Potatoes: The Effect Of Biostimulants And Herbicides