





A common myth suggests that dogs don’t see color, that they exclusively see the world in shades of grey. Dogs have two cone photoreceptors within the retina, compared to humans, who have three. The spectral peaks for these two cones almost identically match up to human short-wave and long-wave sensitivities in humans (spectral peaks at 555 nm and 429 nm).
Recent research by Siniscalchi et al. (2017) suggests that dogs exhibit behavioral responses similar to those of red-green deficient humans on a canine modified Ishihara test (Ishihara, 1918). Dogs oriented less to red-green color contrasts that are more difficult for red-green color-blind humans to differentiate. Tanaka et al. (2000) found that two dogs were able to successfully select a colored stimulus (red, blue, or green) compared to a grey one.
However, in addition to color, dogs may also be using brightness cues to identify and discriminate between stimuli. In a small sample of dogs, Rosengren (1969) found that Cocker Spaniels successfully discriminated red, blue, green, and yellow colored dishes across various levels of brightness. Yet, Kasparson et al. (2013) suggests that dogs preferentially use color cues (when discriminating yellow and blue stimuli) over brightness cues, under natural lighting conditions. Taken together, these previous findings suggest dog color perception is more complex and an essential feature.
What we did
To evaluate color perception in dogs, specifically red and green, and blue and yellow, contrasts presented on a computer monitor, the study published in Applied Animal Behavior Science evaluated seven dogs’ ability to differentiate between various foreground and background colors at different physical luminance levels. The subjects were trained to select one of two circles (either larger or smaller) in a two-choice size discrimination task. The colors of the circle stimuli were then varied so that dogs would choose one stimulus (a colored one) if they could discriminate between two colors, and a different stimulus (a black one) if they could not.
What we found
We conducted four experiments. The first presented dogs with yellow, blue, green, and red stimuli at their maximum red, green, and blue (RGB) values (e.g. 0, 0, 255). All seven dogs were extremely successful at discriminating between red and green, yellow and blue, and black and white stimuli, and no significant difference in performance was observed (p = 0.52). In Experiments 2 to 4, the four colors were presented with equal physical luminance (in other words, brightness) across three different intensities (ranging on a scale from dimmer to brighter). All dogs successfully discriminated between all four colour contrasts when the colors were isoluminant at 34.6 cd/m2 (yes, this measure literally means the number of candles bright in a meter squared of space) as no difference in performance was observed (p = 0.10).
When presented with isoluminant stimuli at 41.9 cd/m2, a difference was observed between conditions. Dogs failed to detect red stimuli presented on a green background compared to the black and white controls (p = 0.03). When presented with stimuli at 49.3 cd/m2 (the brightest presentation of equally bright stimuli), a difference was again observed between conditions. Contrary to expectation, dogs were unable to detect blue stimuli presented on a yellow background compared to the achromatic controls (p < 0.01).
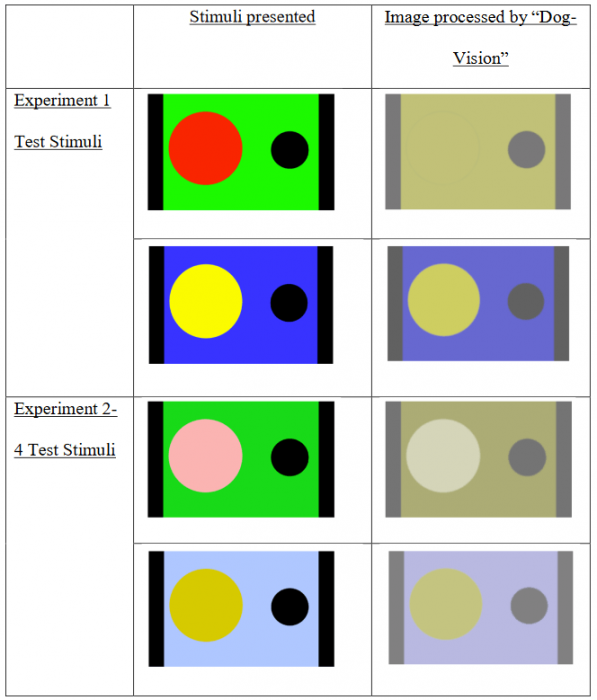
Unaltered and “Dog-Vision” altered stimuli from Experiments 1 to 4. Examples of stimuli from Experiments 1 to 4 in their unaltered form as well as the processed images generated by (Pongrácz et al., 2017). Dogs were capable of identifying the missing red circle in the image processed by “Dog-Vision” in Experiment 1.Contrary to the proposed to be visible circle by the image processing tool, dogs demonstrated difficulty in identifying a blue circle stimulus presented on a yellow background (Experiment 4). Figure courtesy Sarah-Elizabeth Byosiere.
What this means
These findings suggest that luminance levels of stimuli can affect color discrimination capabilities in dogs, particularly when presented on computer screens. These results have critical implications in how cognition is studied in dogs and offer additional insight into tools such as Dog-Vision, that alter images into a proposed depiction of what dogs see. While an interesting concept, the image-processing tool simply calculates color perception of color based on maximum RGB values.
While this image processing tool has been evaluated (Pongrácz et al., 2017), it has only been tested in humans. When humans are presented with images processed by the software, results suggest they have a more difficult time interpreting certain communicative cues. However, it has yet to be validated or assessed in a dog population. Our investigation suggests that the algorithm underlying the “dog-vision” does not line up with actual canine perception as dogs were capable of identifying test stimuli (specifically the red and green contrasts) in Experiment 1, contrary to what the software would predict (see figure 7 in the manuscript).
How to interpret this
While some have proposed that care should be taken when throwing a red ball on green grass for dogs, this is an over exaggeration. There are various other cues — brightness, shape, size, contour, and more — that dogs are likely using to pick up on visual stimuli, even when color may be difficult to identify. Just because they see the world differently does not mean the ball entirely disappears from their visual field. It just means they likely have other ways to identify it. Our results interestingly suggest that even when red-green stimuli are identical in physical brightness, discrimination is still possible. What cues might they be using and how?
As stimuli became increasingly bright across the stimuli increased, certain color contrast discriminations became harder. At 41.9cd/m2 (Experiment 3), dogs generally struggled to discriminate red stimuli presented on a green background but were capable of successfully discriminating the inverse, green stimuli on a red background. This may be due to differences in color contrast discrimination, as performance may be dependent on the organization of the background and foreground color. In humans, color-opponent cells can be excited and inhibited based on the neuronal signal on the center and surround of the receptive field (Hubel and Wiesel, 1959, 1961, 1962, 1965). While neurons may receive identical wavelengths in both the red on green, and green on red conditions, different excitatory and inhibitory processes may occur as different areas of the receptive field are affected.
An unexpected finding was that the dog’s group performance on the blue stimuli presented on a yellow background differed from controls when presented at 49.3 cd/m2 (Experiment 4). As dogs do not physiologically lack short-wave cones, we suggest that this finding may be the result of color saturation. As brightness increases, colors become more saturated and begin to converge as a white hue. It is possible that dogs are particularly prone to saturation effects given that their vision is more scotopic than it is photopic (meaning their visual system is designed to function more optimally in dim light). Therefore, this yellow and blue contrast may have been perceived as two varying shades of white hues.
It is important to note that the stimuli presented in this study were presented at equal physical luminance. One question that has plagued vision and color researchers for centuries is, “Is the red that I see the same as the red that you see?” In humans, we can ask study participants to match up two colors (or brightness’s of one color) until they are indiscernible from one another. This point actually varies for individuals. One subject may just be a little different from the other. This point represents an individual’s perceptual isoluminance. While the stimuli may be physically equal in brightness, a subject point of where that perceptually looks equally bright may differ. This is nearly impossible to study in animals, and therefore one limitation, and likely also a reason for individual variation in the dogs, is that we were only able to look at physical isoluminance and not perceptual isoluminance.
Related Posts
 Genes Keep You On A Long Leash With Your Dog
Genes Keep You On A Long Leash With Your Dog Looking At Versus Focusing On Faces: What Attracts Our Attention?
Looking At Versus Focusing On Faces: What Attracts Our Attention? What Do They Get Out Of It? The Emotional Experience Of Providing Social Support
What Do They Get Out Of It? The Emotional Experience Of Providing Social Support How Does Rewarding Safe Choices Affect Teen Decision Making In Peer Contexts?
How Does Rewarding Safe Choices Affect Teen Decision Making In Peer Contexts? Adolescent Self-Efficacy Is Shaped By Family, School, And Peers
Adolescent Self-Efficacy Is Shaped By Family, School, And Peers Does Morality Make Us Make God In Our Own Image?
Does Morality Make Us Make God In Our Own Image?