





At the time of George Perkins Marsh’s book ‘Man and Nature’, published in 1864, people realized that human activities have an impact on the environment. Since then, ecologists developed new metrics to assess such impacts, in order to understand the changes happening in the field and develop preventive or curative measures.
Most of the metrics to assess anthropogenic impacts used, and still use, features of living communities, such as species richness. More recently, however, ecologists also began focusing on species quality by measuring the diversity of evolutionary lineages within habitats and regions, as well as functional traits of particular ecological importance (e.g., body mass, type of locomotion, specializations of the feeding apparatus etc.). These metrics are commonly used by governmental agencies in different countries when monitoring the quality of the environment and are at the basis of political decisions for environmental regulators.
The general idea is that any impact on an ecosystem will impair some aspect of its communities of organisms, for example by reducing the number of species with local extinctions. Among the best-known cases, in streams and rivers, human-induced eutrophication from nearby farms will increase consumption of oxygen and thus cause a reduction in the presence of those species that have high oxygen demands; similarly, in lakes, the release of planktivorous fish for sport fishing will completely change the size spectra of zooplankton, with a reduction of larger crustaceans; indeed, the list of examples of differential response, depending on ecological features, is long and rather well studied.
Yet, one aspect that is not usually considered in such assessments is related to morphological variability at the community level. An ecological community is a group of species living in a certain geographical region or habitat. As anticipated, communities vary in composition, as well as in the functions of their members. However, they can also differ in the number of types of organismal forms that live in those communities. Thus, we wondered whether, as for species abundance and richness, we might expect fewer differences in size or shape across a group of related organisms in strongly anthropized habitats. In this context, each habitat, or area within a larger region, might be seen as a landscape made of different combinations of morphologies, which, together, make up the local ‘morphoscape’. In fact, the ‘morphoscape’ concept has been previously used, within a single species, in paleontology, but no one has explored its usefulness in ecology to assess the morphological complexity of a community.
In our study, we focused on a widespread and diverse family of beetles, the Carabidae (Fig. 1), and measured how big their morphoscape is across four types of habitats (cropland, meadow, poplar stand, and woodland) found in the Padana Plain of northern Italy. Using museum specimens, and available information on species composition, we selected a single adult male individual for each species found in the four habitats and applied a method of image analysis to quantify body size and body shape using digital photos. Within a habitat, the magnitude of the morphoscape was quantified using different metrics. The most intuitive of these is simply the range of variation, which is analogous to computing the difference between the tallest and shortest person in a group of people to estimate how much individuals differ in that sample.
We discovered that body size partly mirrored differences in species richness, with more variation in the habitat with more species, regardless of human impact. Thus, open habitats, such as meadow and cropland, housed a wide range of body sizes, as well as more species of carabid beetles (respectively ca. 100-110), despite being under stronger anthropogenic pressures. However, woodland, the least impacted habitat, had about as much size variation across species as cropland and meadow, and consistently showed the biggest morphoscape for shape (Fig. 2). This is particularly remarkable if one considers that cropland, in fact, had a morphoscape almost as big as woodland, but woodland reached that magnitude despite being host to almost 20% fewer species than cropland. This suggests that even if a relatively pristine closed woodland habitat could be less species-rich, it might, in fact, conceal a relatively higher number of morphologies and be thus potentially richer in the ecological functions played by its fewer but more disparate members. Using an analogy, it is like having two companies, that we call C and W, renting vehicles for different types of work: both companies have a full range of sizes going from small bikes to big trucks; C has more brands; however, W can offer some less common types of vehicles, for instance, auto-rickshaws and caravans, that are not available at C.
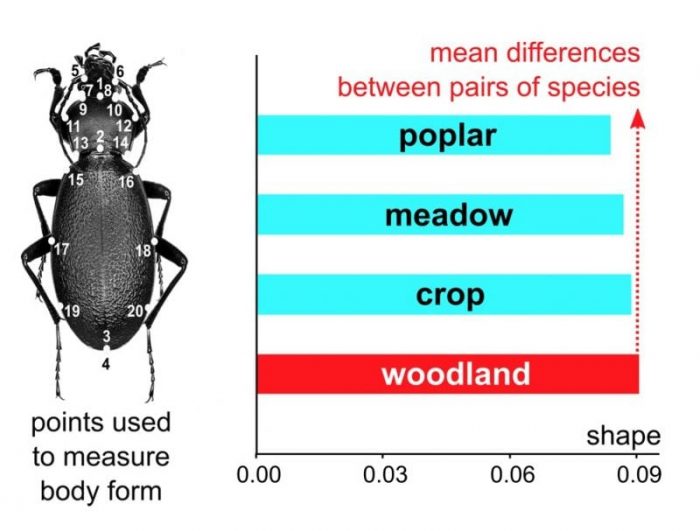
Fig. 2. Configuration of anatomical landmarks used to describe Carabid beetle body form (left) and a bar chart (right) of mean shape variability showing a bigger morphoscape for woodland compared to the other three Padana plain habitats. (Andrea Cardini)
That the biggest morphoscape for carabid body shape in the Padana Plain occurs in the least anthropized habitat raises some intriguing questions. Shape, a highly multivariate and complex trait, often indicative of adaptations, might be informative on both genetic and ecological variability, including some of the ecological functions that might be impacted by human activities. In fact, however, although form and function often correlate, which is what allows anatomists to predict the ecology and behavior of extinct animals such as dinosaurs, for now, we cannot be sure that more disparate shapes necessarily correspond to ecologically more complex habitats. Future studies using similar approaches in beetles from other regions, as well as in other groups of organisms, will be needed to find robust answers. Excitingly, the future might also bring some interesting parallels with other less conventional ecological disciplines, such as sound ecology, in which there is a growing body of evidence showing that healthy ecosystems might indeed have more complex acoustic landscapes or soundscapes.
These findings are described in the article entitled Estimating the magnitude of morphoscapes: how to measure the morphological component of biodiversity in relation to habitats using geometric morphometrics, published in the journal The Science of Nature.
This work was led by Diego Fontaneto (National Research Council of Italy), Martina Panisi (Faculdade de Ciências da Universidade de Lisboa), Mauro Mandrioli (University of Modena and Reggio Emilia), Dario Montardi (University of Modena and Reggio Emilia), Maurizio Pavesi (Museo Civico di Storia Naturale), and Andrea Cardini (Università di Modena e Reggio Emilia).
Related Posts
 Duck-Billed Dinosaurs Uncovered In Aniakchak, Alaska
Duck-Billed Dinosaurs Uncovered In Aniakchak, Alaska Cryptic Diversity In Vietnam’s Limestone Karst Habitats
Cryptic Diversity In Vietnam’s Limestone Karst Habitats An Improved Method To Remove Debris From Cyst Nematode Egg Suspensions And Computer-Aided Technologies For Egg Counting
An Improved Method To Remove Debris From Cyst Nematode Egg Suspensions And Computer-Aided Technologies For Egg Counting The Footprints Of Urbanization, Industrialization, And Agriculture On River Beds: Heavy Metal Contamination Assessment And Source Identification In River Sediments In Eastern China
The Footprints Of Urbanization, Industrialization, And Agriculture On River Beds: Heavy Metal Contamination Assessment And Source Identification In River Sediments In Eastern China Aging Dolphins Via Pectoral Flipper Radiography
Aging Dolphins Via Pectoral Flipper Radiography Glycoalkaloids In Potatoes: The Effect Of Biostimulants And Herbicides
Glycoalkaloids In Potatoes: The Effect Of Biostimulants And Herbicides