





Introduction
Tall stem-rosettes — aka giant rosettes — grow at high elevations on many tropical mountains, including the northern Andes of South America, the East African highlands, and the large volcanoes of the Hawaiian and the Canary Islands. In contrast to trees, the woody trunks of these plants do not generate annual growth rings, thus their age cannot be readily determined; instead, variation in stem height has routinely been used as a proxy for estimating plant age. Elongation rates have been measured for several stem-rosette species during the past 50 years; average growth rates, determined for small populations or groups of stem-rosettes, have then been used to calculate plant age. Difficult access to the mountain areas where these plants grow, and various logistic reasons, have restricted most growth studies to short time periods, often for less than one to just a few years.
Andean stem-rosettes are collectively known in South America as frailejones (singular: frailejón, “big monk”), due to the alleged resemblance of plants to a monk; frailejones are found along the Andean Cordillera over a ≥1200 km distance, from central Ecuador to northern Venezuela (Cuatrecasas, 2013). Research reported here focused on two tall, abundant frailejón species in the Páramo Piedras Blancas, Sierra La Culata, Venezuela, at 8°52’ N. Elongation rates were measured for 76 Coespeletia timotensis and 38 Espeletia schultzii at 4270-4525 meters (~14010-14850 feet) elevation, for periods ranging between 10 and ~20 years; plants were relocated with the help of repeat photography. In this mountain range, C. timotensis reaches its maximum altitude at ~4565 m (~14980’), although in the nearby Sierra Nevada de Mérida it is found at 4780 m (~15685’); actually, this plant grows at the highest elevation reported for any of the 144 frailejón species in the Andes (Diazgranados, 2012). E. schultzii, the other plant studied, is also common in this range, up to ~4300 meters (~14110’).
Climatic context and growth of stem-rosettes
Harsh climatic conditions at upper elevations severely restrict the development of all plants, including frailejones, which grow slowly. Freezing temperatures occur nearly every night, causing much ground frost, which kills the majority of seedlings; the lower permafrost limit — where mean soil temperature drops below 0°C — occurs in Piedras Blancas at ~4550-4600 m (Schubert, 1975). Extreme soil drought also develops during the 4-month long dry season, further stressing plants. In the northern Andes, the upper boundary for plants is a cold and dry limit, caused by heat deficit and physiological aridity. In spite of the difficult high-mountain conditions, some frailejones grow to a prodigious size; C. timotensis can often attain ~3.5 m-height (Photos). E. schultzii is much shorter and reaches only to ~1.5 m stature.


These photos show a group of C. timotensis at ~4385 m; the one on the right is one of the tallest rosettes I ever found in Piedras Blancas. On Jan. 10, 1982, height to its uppermost leaf tip was 291±2 cm (photo a); on Feb. 29, 2000, after an 18-yr, ~2-mo period, it had grown 21 cm to 312±2 cm (photo b). Using a specific growth rate of ~1.16 cm/yr (see discussion, below) would yield a presumed age of ~269 years for this plant; of course, this would assume, probably incorrectly, a uniform growth rate during this plant’s life. The second tallest rosette to the left seems to have grown even more (~28 cm, ~1.54 cm/yr) during the same time interval; scale is 180 cm-long.
Earlier surveys over brief time spans could not, perforce, detect much appreciable variability in rates of expansion among slow-growing individuals, but assessing elongation for one to two decades allowed small variations to be maximized, and growth differences observed between plants were as high as ~34 cm. Previous average growth for C. timotensis populations was estimated at 1.5 cm/yr, and 1.0 cm/yr for E. schultzii (Smith, 1981). Mean trunk elongation rates during this long-term study were not significantly different: 1.24 ± 0.46 cm/yr, and 1.04 ± 0.40 cm/yr, respectively. However, one surprising and entirely unexpected result was that these average rates concealed a broad variation among individual plants of either species.
Probable causes of variation in plant growth
Even plants growing near each other, occupying similar topographic positions and soil types, and with the same aspect, displayed extreme variability. The graph indicates that stem growth rates (cm/yr) during a 10-yr, ~2-month period for 60 surviving, adjacent C. timotensis along a valley floor at ~4285 m fluctuated between ~0.4 and 2.5 cm/yr. A comparable scattergram for the 31 remaining E. schultzii showed growth rates varied from ~0.4 to 2.4 cm/yr. Significantly, this graph also indicates that the initial plant size was not associated with average growth, which seemed random and unrelated to plant age.
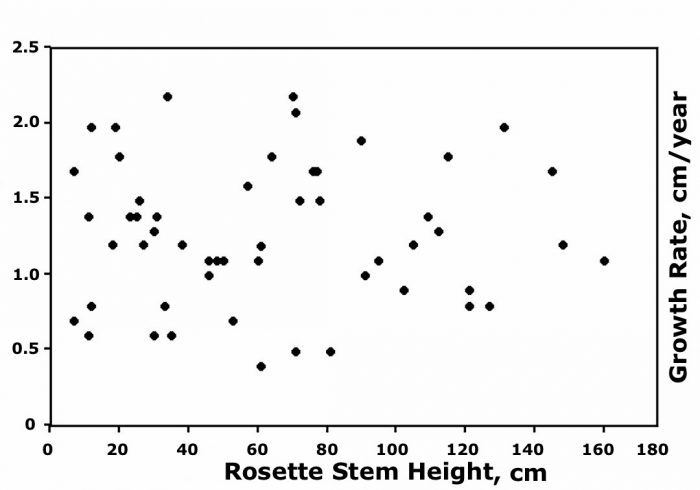
Figure courtesy Francisco L. Pérez.
Conceivably, variation among individual plants could be caused either by differences in physical microsite features — which may change over short distances — or in biological conditions near specific plants. Microsite factors known to affect frailejón growth include soil moisture content, level of soil nutrients, presence of rocks nearby, and competitive interactions between adjacent plants.
Plants may affect the growth of individual stem-rosettes in two ways. Frailejón development can be facilitated by germination on nurse cushion plants, or next to rocks; both reduce frost frequency and disturbance and generally improve soil moisture conditions. Other interactions might inhibit growth through competition; for example, studies of desert columnar cacti have found that vegetation abundance in the immediate vicinity of plants produces an “interference factor” that negatively affects growth rate. Competition was not analyzed in this study, but previous data on páramo vegetation strongly suggest the possibility of plant interference for both species, as the C. timotensis stand had abundant vegetation, with 64% plant cover and 37 vascular plant species; the E. schultzii stand was also densely vegetated, with 45 plant species and 61.5% plant cover.
Plant attrition by mortality
Enough plants survived by the end of the study period to perform critical statistical analyses, but plant mortality reduced both frailejón populations by ~18-19%. C. timotensis seedlings were presumably killed by frost heaving and/or browsing by semiferal cattle. Tall, mature C. timotensis were decimated by plant toppling, a common event for this species; this was probably associated with slope movement and ground shifting due to frost disturbance. Soil instability also affects tall Senecio keniodendron stem-rosettes on Mount Kenya (Africa), where tilted “drunken stands” occur on steep, disturbed slopes (Mahaney, 1980). Due to significant leaf pubescence and high content of resinous compounds, cattle avoid eating E. schultzii; also, and presumably due to their shorter stature, these frailejones were not substantially affected by tilting, and most dead plants remained standing without toppling.
Conclusions
In conclusion, as stem-rosette growth rates, even of adjacent plants, vary by a factor of 6-16, using an average rate to date individual plants seems meaningless; lacking specific growth data for particular plants, this approach seems unwise and ought to be discontinued in future studies. In any case, emphasis on precise dating of páramo frailejones might just be secondary. The most remarkable aspect of these long-lived plants could be how strikingly adapted to the hostile high Andean environment they are, and how they have evolved to occupy a broad range of mountain habitats. C. timotensis, growing at greater elevations than any other Andean frailejón, may well occupy the most prominent evolutionary pinnacle of high Andean plants (Cuatrecasas, 2013).
References:
- Cuatrecasas, J., 2013. A systematic study of the Subtribe Espeletiinae (Heliantheae, Asteraceae).
Memoirs New York Botanical Garden, 107, 1-689. - Diazgranados, M., 2012. A nomenclator for the frailejones (Espeletiinae Cuatrec., Asteraceae).
Phytokeys, 16, 1-52. https://phytokeys.pensoft.net/articles.php?id=1436. www.phytokeys.com. - Mahaney, W.C., 1980. Late quaternary rock glaciers, Mount Kenya, Kenya. Journal Glaciology,
25, 492-497. - Schubert, C. 1975. Glaciation and periglacial geomorphology in the northwestern Venezuelan
Andes. Eiszeitalter und Gegenwart, 26, 196-211. - Smith, A.P., 1981. Growth and population dynamics of Espeletia (Compositae) in the Venezuelan Andes. Smithsonian Contributions to Botany 48, 1-45.
Related Posts
 Duck-Billed Dinosaurs Uncovered In Aniakchak, Alaska
Duck-Billed Dinosaurs Uncovered In Aniakchak, Alaska Cryptic Diversity In Vietnam’s Limestone Karst Habitats
Cryptic Diversity In Vietnam’s Limestone Karst Habitats An Improved Method To Remove Debris From Cyst Nematode Egg Suspensions And Computer-Aided Technologies For Egg Counting
An Improved Method To Remove Debris From Cyst Nematode Egg Suspensions And Computer-Aided Technologies For Egg Counting The Footprints Of Urbanization, Industrialization, And Agriculture On River Beds: Heavy Metal Contamination Assessment And Source Identification In River Sediments In Eastern China
The Footprints Of Urbanization, Industrialization, And Agriculture On River Beds: Heavy Metal Contamination Assessment And Source Identification In River Sediments In Eastern China Aging Dolphins Via Pectoral Flipper Radiography
Aging Dolphins Via Pectoral Flipper Radiography Glycoalkaloids In Potatoes: The Effect Of Biostimulants And Herbicides
Glycoalkaloids In Potatoes: The Effect Of Biostimulants And Herbicides