




Faced with sudden external physical stress, e.g.temperature shifts, desiccation, pH changes, cells need to adapt rapidly. Part of the adaptation is to counter the cellular impact of the stress itself, part is to balance the drop in energy production with a corresponding reduction of energy spending. These changes have to go into effect rapidly and to manage this, the cell needs to operate on multiple fronts simultaneously.
Besides altering the transcription program, pre-existing transcripts can be modulated for increased or decreased turnover rate, and for increased or decreased efficiency of translation into proteins. These regulatory steps taking place at the level of the mRNA are called post-transcriptional. Extensive such regulation is well-known to occur as an important part of the stress response. Much of post-transcriptional regulation is mediated through RNA-binding proteins (RBPs). This has been confirmed mostly in studies of post-transcriptional regulation that were done globally, i.e.considering all transcripts collectively.
We wanted to look at interactions between individual mRNA species and RBPs. Our idea was that stress-induced mRNAs might recruit a different composition of RBPs than most mRNAs, and that this could determine their fate in the stress response. To achieve this, we used the unicellular organism Saccharomyces cerevisiae, a classical model for eukaryotic cells. We modified a previously described procedure for genetic modification to add an affinity tag to the mRNA of interest.
Our main modification was to use a higher affinity GFP-binding protein. This allowed us to use highly specific binding to a solid phase to purify the tagged mRNA away from bulk mRNA. We chose two stress-induced mRNAs and two mRNAs unaffected by stress for tagging and purification. From cells exposed to osmotic stress, we then purified these four tagged mRNAs, each with its own complex of associated proteins. The proteins were analyzed and quantitated with mass spectrometry, and the composition compared between the four mRNAs.
Credit: Per Sunnerhagen
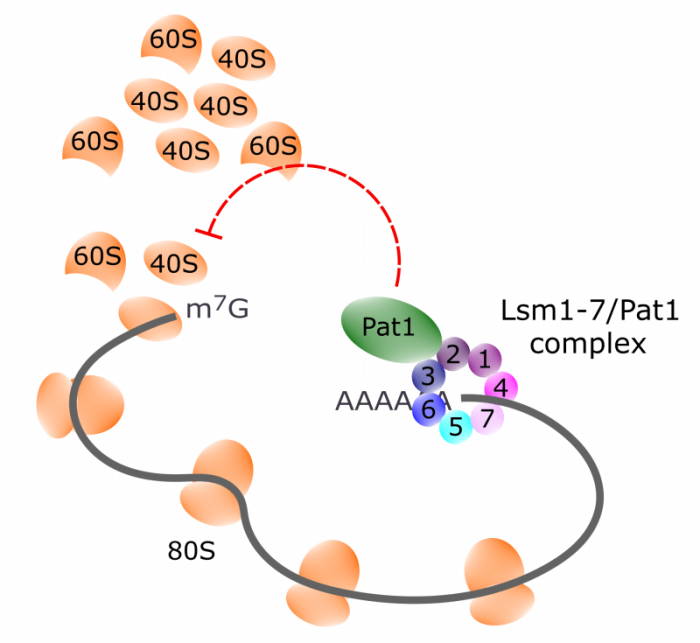
It emerged that proteins from the Lsm1-7/Pat1 complex were enriched in the samples with stress-induced mRNAs, relative to the mRNAs not affected by stress. This complex has been described to bind mRNAs to connect them to the degradation machinery, which competes with the translation machinery. The complex exists in two varieties – the cytoplasmic Lsm 1 ‑ 7 / Pat1 complex, and the nuclear Lsm 2 – 8 / Pat1 complex. These differ only by two subunits; Lsm1 which is present only in the cytoplasmic complex, and Lsm8 which is unique to the nuclear complex. We did recover Lsm1 but not Lsm8, and so concluded that it is the cytoplasmic version of the complex we are dealing with. This is an indication that any effect on the behavior of stress-induced mRNAs exerted by the complex should take place in the cytoplasm. This means there are two main candidates: an effect on translational efficiency, and an effect on mRNA turnover rate.
To see what effects the Lsm1-7/Pat1 complex would have on stress-induced mRNAs, we turned to lsm1and pat1mutants and compared their behavior to wild-type. We found that throughout all phenotypes studied, we found virtually identical results between the two mutants. This informs us that the Lsm1 and Pat1 proteins participate in the same roles, likely as parts of the same protein complex.
Our most remarkable finding was the effect of lsm1 or pat1 mutations on the production of stress proteins. In normal cells, stress proteins are not made unless the stress intensity exceeds a certain limit. Over that limit, the amount of output stress protein depends on the stress level. In lsm1 or pat1 mutants, however, even a tiny stress level was enough to provoke production of stress proteins at near-maximal levels. You could say that the mutant cells had become trigger happy about their stress response. The interpretation of this is that the Lsm 1-7/Pat1 complex serves to dampen the stress response, preventing it from firing when it is not needed.
We then compared the mutant-specific increases in protein and mRNA levels, respectively, and calculated that the protein increase was too large for this to be accounted for by the mRNA increase alone. This was a pointer that this effect was due to an impact of Lsm1‑7/Pat1 on translation. To investigate this possibility, we turned to a recently developed method to trace the passage of ribosomes over the mRNA molecule during translation. The method relies on the finding that most mRNA molecules that have started to degrade from their 5’-end do this because they are in the process of being translated by ribosomes, which entered by the 5’-end and are moving towards the 3’-end of the mRNA. This means that by considering only the mRNA molecules that lack the “cap” (a specialized structure at the 5’-end, which is removed while degrading from this end), and counting the number of molecules with their 5’-end at a particular nucleotide, we get a quantitative picture of the position of the ribosome on that mRNA while it was being translated. One can thus monitor the speed of transit of the ribosome over different parts of the mRNA.
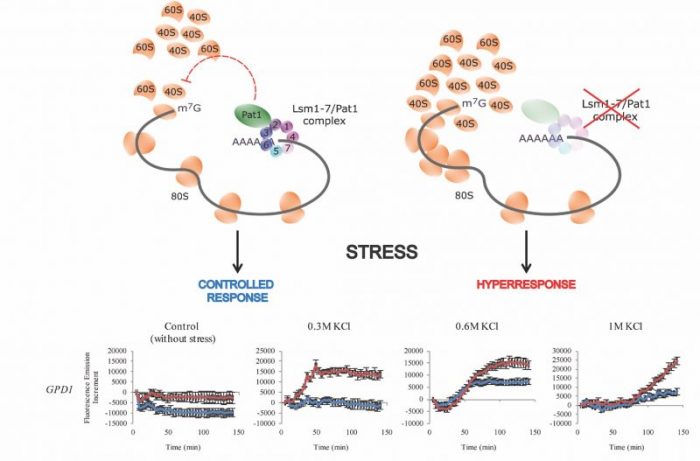
The Lsm1-7/Pat1 complex is associated with the 3’ end of the mRNA (left). Through interaction with the 5’ end, it limits the access of ribosomes to start translation. This leads to the controlled production of stress protein (blue lines in lower diagrams). If Lsm1-7/Pat1 is missing (right), ribosomes have increased access to start translation of stress proteins, and hyper-response results (red lines in lower diagrams). Republished from Open Access journal PLOS from https://doi.org/10.1371/journal.pgen.1007563.
We noticed that in the lsm1 and pat1 mutants, there was an increased accumulation of mRNA 5’-ends in the region just before the translation start (the 5’-untranslated region). The accumulation was further accentuated for stress-activated transcripts. This means an increased buildup of ribosomes in this region in the mutants. In light of the exaggerated production of stress-activated proteins in the mutants, we interpreted this as an increased accessibility to the 5’-untranslated regions for ribosomes. Thus, one function of the Lsm1-7/Pat1 complex may be to diminish the access of ribosomes to this mRNA region, and so to limit the stress response to conditions where it is really needed.
This protein complex is best known for its activities at the 3’-end of mRNAs, where it is involved in regulating the turnover rate of the transcript. However, there are recent reports suggesting that it also has a role in regulating translation. Our results from 5’-phosphate sequencing rather pointed at a role at the other end of the mRNA. These ideas are not mutually exclusive. During active translation, an mRNA forms a loop where its two ends are brought in physical proximity by bridging RNA-binding proteins. The Lsm1-7/Pat1 complex is evolutionarily old and conserved among all eukaryotes, so there is reason to assume that it will work in similar ways in plants or animals, including humans. Our study supports the notion that one of its functions is to provide the right amount of stability to the expression system and prevent undue release of stress responses.
These findings are described in the article entitled The Lsm1-7/Pat1 complex binds to stress-activated mRNAs and modulates the response to hyperosmotic shock, recently published in the journal PLOS Genetics. This work was conducted by Elena Garre and Per Sunnerhagen from the University of Gothenburg, Vincent Pelechano from the Karolinska Institute, and Manuel Sanchez del Pino and Paula Alepuz from the Universitat de Valencia.
Related Posts
 Important Strategies To Help Healthcare Providers Support Patients With Diabetes
Important Strategies To Help Healthcare Providers Support Patients With Diabetes Studying The Link Between Increased BMI And Late-Onset Preeclampsia In Pregnant Women
Studying The Link Between Increased BMI And Late-Onset Preeclampsia In Pregnant Women A Bacterial Cell Imaging Method Using CRISPR And Microfluidics
A Bacterial Cell Imaging Method Using CRISPR And Microfluidics B. infantis Reduces Key Markers Of Intestinal Inflammation In Infants
B. infantis Reduces Key Markers Of Intestinal Inflammation In Infants Epigenetic Changes In Multiple Sclerosis – Studied In Twins
Epigenetic Changes In Multiple Sclerosis – Studied In Twins Did You Know Math Can Help Us Learn How Diseases Work?
Did You Know Math Can Help Us Learn How Diseases Work?