





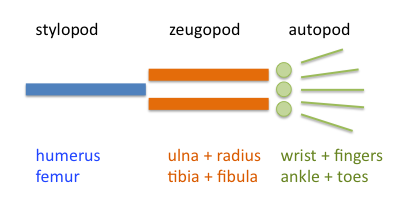
Limbs can take many forms, from paws to wings, allowing vertebrates to walk, run, swim, fly, and grab things. And even though limbs look very different on the outside, they all share a common anatomical design that evolved from the lobe-fins of water-dwelling vertebrates about 370 million years ago. Compelling evidence from fossils, embryology, and comparative anatomy supports this major evolutionary transformation. However, there are still some unknowns about the evolution of limbs; in part, because early forms show a mosaic of features between fins and limbs and some parts are not homologous.
{kind=link}
In a recent study published in Science Advances, we tried to understand better the fins-to-limbs transition by examining what anatomical changes fins went through to become limbs, and how similar these changes were in pectoral and pelvic fins and limbs. We have included in this study some of the most iconic extinct early tetrapods (4-legged vertebrates), such as Tiktaalik, Acanthostega, or Ichthyostega; as well as living fossils, such as the coelacanth fish and the tuatara (Figure 1).
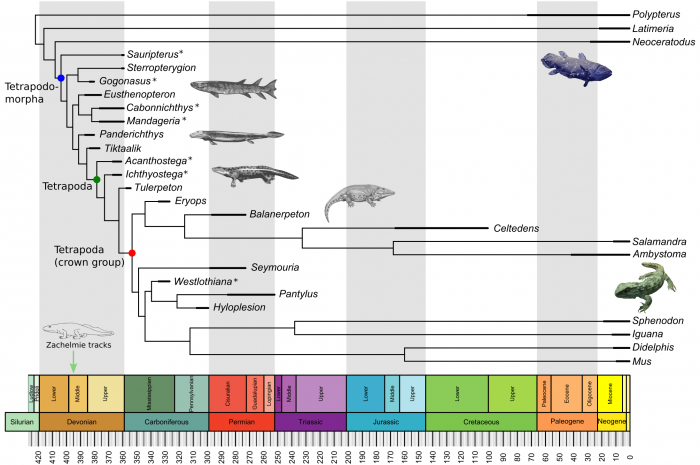
Figure 1. The evolutionary context of the fins-to-limbs transition with the species included in the study. For some species (*) only the complete anatomy of the pectoral or the pelvic appendage is known. Figure courtesy Borja Esteve-Altava.
In the first part of the study, we created network models of the whole skeleton of fins and limbs of modern and extinct vertebrates spanning the fins-to-limbs transition. In these models, each node represents one bone and each link represents one physical joint between a pair of bones. We quantified and compared the anatomy of fins and limbs using network parameters calculated on these models (Figure 2). Traditionally, researchers often take two different approaches when comparing very different forms as fins and limbs: either they compare qualitatively the whole anatomy (as in comparative anatomy) or they compare quantitatively the few elements that we know are homologous (using morphometrics). Modeling fins and limbs as networks of connected bones has allowed us to quantify and compare the anatomy of evolutionary related, but very different, structures in a way that is not accessible to other current methods for measuring anatomy.
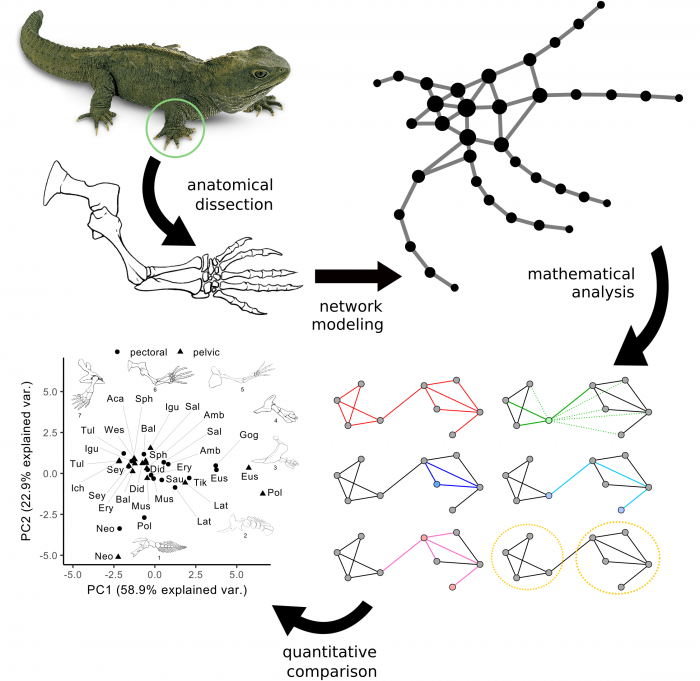
Figure 2. General conceptual approach of anatomical network analysis in our study. Fins and limbs are first modeled as networks, for which we calculated a set of parameters that we can use to compare them with each other. Figure courtesy Borja Esteve-Altava.
In the second part of the study, we used the network parameters measured on fins and limbs to answer questions about the evolution of their anatomy using phylogenetic comparative methods. Namely, (1) what is the main difference between fins and limbs (e.g. knee/elbow or fingers/toes)? (2) How similar fins and limbs were in the past and how they changed over time? (3) Did pectoral and pelvic appendages change in parallel or each one on their own? And (4) did the similarity between pectoral and pelvic appendages increased or decreased over time since the origin of tetrapods?
We asked whether there are differences between appendages with and without digits (fingers and toes), between pectoral and pelvic appendages, and between extinct and modern species. To answer that we combined all network parameters (using a principal components analysis) and assessed statistically the differences between these groups.
We found that having digits is enough to clearly discriminated limbs from fins, due to the resulting morphological changes (Figure 3). These changes include having bones that are less connected in general but more heterogeneously connected (some have many articulations, some have very few), having a greater assortativity (bones with a similar number of articulations contact to each other), and being more modular (limbs divide into blocks of inter-connected bones). However, we found no difference for the other hypotheses tested (pectoral vs pelvic; fossil vs modern). The finding of higher modularity in limbs compared to fins is particularly interesting because modularity has been related to the potential to diversify and acquire new specializations, as limbs did.
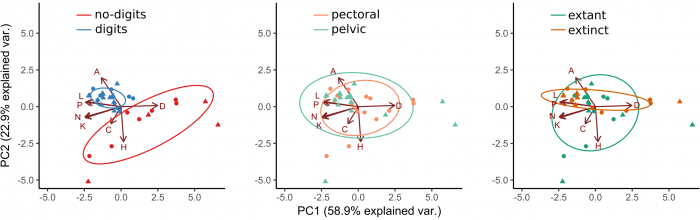
Figure 3. Discrimination of types of appendages. Figure courtesy Borja Esteve-Altava.
Then, we evaluated whether appendages were more similar early in evolution than later on. To answer this question we performed what is known as a disparity through time analysis, in which the overall similarity of all the appendages present at a given time is calculated and compared to that expected by chance. We found that disparity decreased for both appendages likewise (Figure 4). Meaning that existing appendages were more similar among them as they evolved across the fins-to-limbs transition and into modern forms. This similarity became more marked at a time concomitant with the earliest-known tetrapods footprints (the Zachełmie tracks), precisely when vertebrates set foot on the ground for the first time.
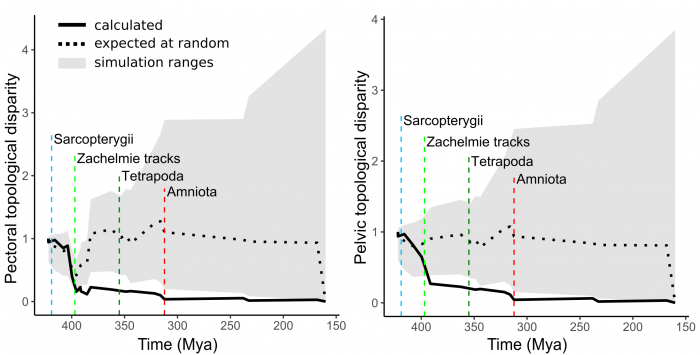
Figure 4. Disparity through time series showed a drop in the amount of differences (i.e. increase of similarity) among the appendages present at that moment in time. Figure courtesy Borja Esteve-Altava.
When looking for convergent evolution of pectoral and pelvic appendages we saw no difference between lobe-fins and limbs, but a common evolution and differentiation of the appendages of sarcopterygians (a group that includes tetrapods, lobe-finned fishes, and their extinct relatives) from those of other fishes (Figure 5). We acknowledge that our small sample may impose some limits on this particular comparison, so we look forward to the discoveries of new fossil materials that allow us to improve these tests. However, the combined findings of the principal component analysis discrimination, disparity through time, and convergence clearly point to parallel evolution of pectoral and pelvic appendages. But, did their similarity within the same animal also change during and after the fins-to-limbs transition?
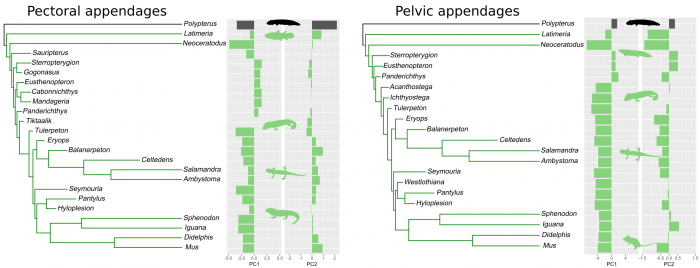
Figure 5. Evolution of convergences in pectoral and pelvic appendages (green indicate convergent evolution). Figure courtesy Borja Esteve-Altava.
To answer this last question we designed a statistical test for how similar the pectoral and pelvic appendages of each species are against the time of the species’ origin (Figure 6). For each parameter, we calculated how much their pectoral-pelvic similarity deviated from a perfect match (absolute residuals on a 1:1 line). We compared these values to how far in time these species lived from the origin of tetrapods. We saw no trend in this relationship, meaning that there was no increase or decrease of similarity after this point in time, and hence, no bottleneck. This finding contradicts some other studies, based on the number of skeletal and muscular elements, that found a similarity bottleneck during this period of time.
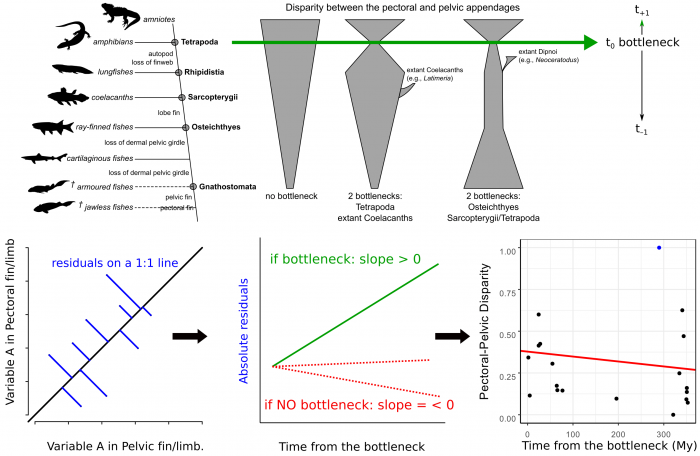
Figure 6. Design of the evaluation of the bottleneck hypothesis. Figure courtesy Borja Esteve-Altava.
In conclusion, our study shows that pectoral and pelvic appendages evolved in parallel during the fins-to-limbs transition, by developing anatomically similar limb designs (they were not so similar before that) and increasing their similarity over time. We interpreted these findings as a compromise between different functional demands and shared developmental constraints in pectoral versus pelvic appendages during the water-to-land transition.
These findings are described in the article entitled Evolutionary parallelisms of pectoral and pelvic network-anatomy from fins to limbs, recently published in the journal Science Advances.
Related Posts
 Duck-Billed Dinosaurs Uncovered In Aniakchak, Alaska
Duck-Billed Dinosaurs Uncovered In Aniakchak, Alaska Cryptic Diversity In Vietnam’s Limestone Karst Habitats
Cryptic Diversity In Vietnam’s Limestone Karst Habitats An Improved Method To Remove Debris From Cyst Nematode Egg Suspensions And Computer-Aided Technologies For Egg Counting
An Improved Method To Remove Debris From Cyst Nematode Egg Suspensions And Computer-Aided Technologies For Egg Counting The Footprints Of Urbanization, Industrialization, And Agriculture On River Beds: Heavy Metal Contamination Assessment And Source Identification In River Sediments In Eastern China
The Footprints Of Urbanization, Industrialization, And Agriculture On River Beds: Heavy Metal Contamination Assessment And Source Identification In River Sediments In Eastern China Aging Dolphins Via Pectoral Flipper Radiography
Aging Dolphins Via Pectoral Flipper Radiography Glycoalkaloids In Potatoes: The Effect Of Biostimulants And Herbicides
Glycoalkaloids In Potatoes: The Effect Of Biostimulants And Herbicides